- Original Paper
- Published:
Epac1 deficiency inhibits basic fibroblast growth factor-mediated vascular smooth muscle cell migration
The Journal of Physiological Sciences volume 69, pages 175–184 (2019)
Abstract
Vascular smooth muscle cell (VSMC) migration and the subsequent intimal thickening play roles in vascular restenosis. We previously reported that an exchange protein activated by cAMP 1 (Epac1) promotes platelet-derived growth factor (PDGF)-induced VSMC migration and intimal thickening. Because basic fibroblast growth factor (bFGF) also plays a pivotal role in restenosis, we examined whether Epac1 was involved in bFGF-mediated VSMC migration. bFGF-induced lamellipodia formation and migration were significantly decreased in VSMCs obtained from Epac1−/− mice compared to those in Epac1+/+-VSMCs. The bFGF-induced phosphorylation of Akt and glycogen synthase kinase 3β (GSK3β), which play a role in bFGF-induced cell migration, was attenuated in Epac1−/−-VSMCs. Intimal thickening induced by the insertion of a large wire was attenuated in Epac1−/− mice, and was accompanied by the decreased phosphorylation of GSK3β. These data suggest that Epac1 deficiency attenuates bFGF-induced VSMC migration, possibly via Akt/GSK3β pathways.
Introduction
Intimal thickening that occurs after vascular injury resulting from percutaneous coronary intervention is a major clinical problem [1]. This thickening is the fundamental process of wound healing in which the vascular smooth muscle cells (VSMCs) migrate toward the internal lumen and proliferate within the innermost layer of the arterial wall [2]. Although drug-eluting stents significantly reduce intimal thickening, repeat revascularization remains relatively common [3], and the risk of definite late stent thrombosis has emerged as a major concern [4]. It is thus important to elucidate the molecular mechanisms involved in the pathological process of intimal thickening.
VSMC migration is a critical event in restenosis [2, 5], and is mediated by the cytokines and growth factors released from the arterial wall [6, 7]. Basic fibroblast growth factor (bFGF) and platelet-derived growth factor (PDGF)-BB are released by endothelial cells, macrophages, and VSMCs at the vascular injury site [8]. Both bFGF and PDGF-BB contribute significantly to VSMC migration and the subsequent intimal thickening [6, 9].
We previously demonstrated that an exchange protein activated by cAMP (Epac) induced the migration of VSMCs and other cell types [10,11,12]. When Epac1 was silenced, PDGF-BB-induced VSMC migration and mechanical injury-induced intimal thickening were attenuated [13]. Another line of study reported that genetic knockdown of the Epac1 gene reduced intimal thickening in a mouse carotid artery ligation model [14]. The contribution of Epac1 to bFGF-induced VSMC migration and intimal thickening, however, remains unknown.
bFGF activates the phosphoinositide 3-kinase (PI3 K)/protein kinase B (Akt) pathway and promotes cell migration [15,16,17,18]. PI3 K/Akt signaling was activated after balloon catheter injury of rat carotid arteries [19]. Epac also stimulates PI3 K/Akt signaling pathways in a variety of cell types [20,21,22,23]. In Epac1-deficient VSMCs, the protein expression of the PI3 K p110 isoform and the phosphorylation of Akt at Ser473 was significantly decreased [14]. These findings suggest that bFGF and Epac share downstream signaling pathways, namely, PI3 K/Akt. We therefore hypothesized that the downstream signaling of Epac1 is involved in bFGF-induced VSMC migration and neointima formation. Here, we demonstrate the effect of Epac1 deficiency on bFGF-induced migration using a primary culture of VSMCs from Epac1−/− (Epac1−/−-VSMCs).
Method
Reagents
Anti-FGFR1 and anti-phosphorylated cofilin antibodies were purchased from Santa Cruz Biotechnology (San Diego, CA, USA). Anti-Forkhead box transcription factor O1 (FOXO1) antibody was purchased from abcam (Cambridge, MA). Anti-glycogen synthase kinase 3β (GSK3β), anti-phosphorylated GSK3β (Ser9), anti-Akt, anti-phosphorylated Akt (Ser473), anti-p70 ribosomal protein S6 kinase (S6 K), anti-phosphorylated S6 K (Thr389), and anti-phosphorylated FOXO1 (Ser256) antibodies were purchased from Cell Signaling (Beverly, MA, USA). Hoechst 33342, rhodamine conjugated anti-F-actin antibody, Alexa Fluor 488 anti-rabbit IgG antibody, and Alexa Fluor 594 anti-rabbit IgG antibody were purchased from Life Technologies (Carlsbad, CA, USA). Recombinant human bFGF protein was purchased from Thermo Fisher Scientific (San Jose, CA, USA). Poly-l-lysine was purchased from Sigma (St. Louis, MO, USA). Elastase type II-A, trypsin inhibitor type I-S, bovine serum albumin V, penicillin–streptomycin solution, and Dulbecco’s modified Eagle’s medium (DMEM) were purchased from Sigma-Aldrich. Collagenase type II was purchased from Worthington Biochemical (Lakewood, NJ, USA). Collagenase dispase was purchased from Roche Diagnostics (Tokyo, Japan). Fetal bovine serum (FBS) was purchased from Equitech-Bio (Kerrville, TX, USA). Tissue-Tek OCT was purchased from Sakura Finetek (Torrance, CA, USA).
Animals
All animals were cared for in compliance with the guiding principles of the American Physiological Society. The experiments were approved by the Ethical Committee of Animal Experiments at Yokohama City University (reference number: F-A-14-108). The generation of Epac1-deficient mice was described previously [24]. Epac1+/+ and Epac1−/− littermate mice (3–8 months old) were obtained from heterozygote crosses.
Isolation and culture of mouse VSMCs
VSMCs were isolated from the thoracic aorta of mice using an explant method as previously described [13]. Briefly, after mice were euthanized with 100 mg/kg of pentobarbital (Kyoritsu Seiyaku, Tokyo, Japan), the aortae were isolated. The aortae were incubated with a collagenase enzyme mixture that included elastase type II-A, trypsin inhibitor type I-S, bovine serum albumin V, collagenase type II, and collagenase dispase, and the tunica media was separated from the adventitia. The tunica media was then cut into small pieces and plated on a poly-l-lysine-coated culture dish. VSMCs were grown in DMEM supplemented with 20% FBS and 1% penicillin–streptomycin and in air supplemented with 5% CO2 at 37 °C. Cells within 10 passages were used in the experiments.
Cell migration assay
Cell migration was assessed by tracking the path length as previously described [13]. After VSMCs were serum starved for 48 h, cells were treated with or without 10 ng/ml of bFGF for 8 h. Path lengths of VSMCs were measured using time-lapse microscopy (TE2000 Eclipse, Nikon, Tokyo, Japan). Images were recorded at 20-min intervals during the tracking, and the nucleus of each cell was manually traced for each frame. We performed three independent experiments and found statistically similar results.
Immunoblotting
VSMCs were lysed and sonicated in a lysis buffer containing 150 mmol/l Na2CO3 [pH 11.0] and 1 mmol/l EDTA. Protein expression in whole cell lysates was analyzed by immunoblotting as described previously [13].
Immunocytochemistry
Immunocytochemistry was performed as previously described [13]. Briefly, VSMCs cultured on 12-mm glass coverslips were serum starved for 48 h and then stimulated for 30 min with DMEM alone (control) or bFGF (10 ng/ml). Fixed cells were incubated with primary antibodies overnight at 4 °C. DNA was stained with Hoechst solution. Morphometric analysis was performed using Nikon TE2000-E (Tokyo, Japan).
Femoral artery injury model
Transluminal mechanical injury of the femoral artery was induced by the insertion of a large wire (0.38 mm in diameter, C-SF-15-15, Cook, Bloomington, IN, USA) as previously described [13]. The arteries were harvested 2 weeks after injury and were embedded in OCT compound and snap-frozen in liquid nitrogen. Frozen sections were stored at − 80 °C until use.
Immunohistochemistry
Immunohistochemistry was performed as previously described [13]. Frozen tissue sections were incubated with a primary antibody overnight at 4 °C. The slides were washed 3 times with Tween 20/PBS for 5 min and then incubated with a secondary antibody for 1 h. After six washes with Tween 20/PBS, the DNA was stained with Hoechst solution.
Statistical analysis
Data are expressed as mean ± standard error of the mean (SEM). Statistical analysis was performed using unpaired Student’s t-test in Fig. 1c. A two-way analysis of variance (ANOVA) followed by Bonferroni’s multiple comparison test was used for Figs. 1b, 2b, 3b, d, 4b, d. A value of p < 0.05 was considered significant.
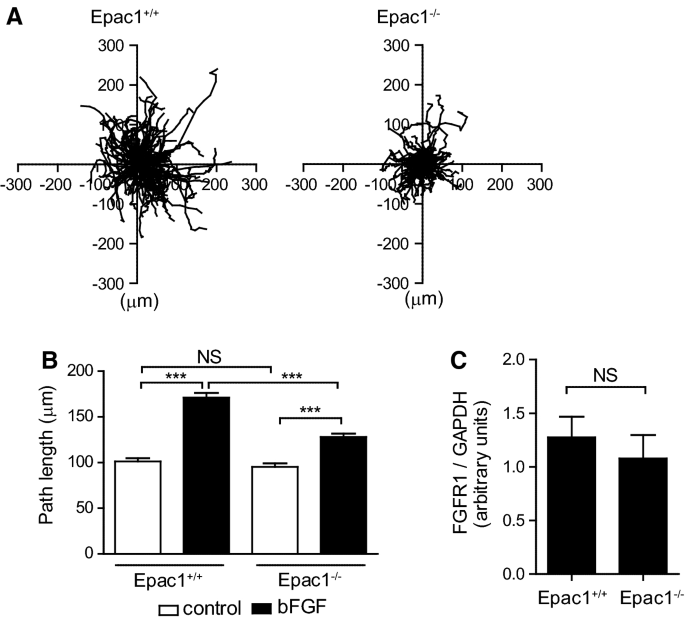
The effect of Epac1 deficiency on bFGF-induced VSMC migration. A Analyses of VSMC migration under stimulation with bFGF (10 ng/ml) for 8 h. Trajectories of VSMC migration from the origin (center of the graphs) to the endpoint. We performed three independent experiments (Epac1+/+-VSMCs, n = 20, 71, and 55; Epac1−/−-VSMCs, n = 23, 40, and 64) and found similar results among experiments. The sum of all data is shown (Epac1+/+-VSMCs, n = 146; Epac1−/−-VSMCs, n = 127). B Quantification of total path length of VSMCs. Data were obtained from three independent experiments (Control Epac1+/+-VSMCs, n = 61, 135, and 65; Epac1+/+-VSMCs treated with bFGF, n = 73, 129, and 46; Control Epac1−/−-VSMCs, n = 26, 126, and 41; Epac1−/−-VSMCs treated with bFGF, n = 40, 105, and 21). Statistical analysis using a two-way ANOVA followed by Bonferroni’s multiple comparison test indicated that all experiments had statistically similar results. The sum of all data is shown (Control Epac1+/+-VSMCs, n = 261; Epac1+/+-VSMCs treated with bFGF, n = 248; Control Epac1−/−-VSMCs, n = 193; Epac1−/−-VSMCs treated with bFGF, n = 166). ***p < 0.0001. NS indicates not significant. C Quantifications of FGFR1 protein expression in unstimulated VSMCs by western blotting. n = 5. NS indicates not significant

Epac1 deficiency decreased bFGF-induced lamellipodia formation. A Representative images of lamellipodia formation in VSMCs. Upper panels: F-actin (red); middle panels: phosphorylated cofilin (p-cofilin, green); lower panels: merged images of F-actin and p-cofilin staining. Arrowheads indicate lamellipodia accompanied by dephosphorylated cofilin. Scale bars, 30 μm. B Quantification of the number of VSMCs with lamellipodia accompanied by dephosphorylated cofilin. Lamellipodia-positive cell rates are presented as the percent of total cell number. n = 353 (control Epac1+/+-VSMCs), 196 (Epac1+/+-VSMCs treated with bFGF), 195 (control Epac1−/−-VSMCs), and 463 (Epac1−/−-VSMCs treated with bFGF). Data were obtained from more than four independent experiments. **p < 0.001; ***p < 0.0001. NS indicates not significant
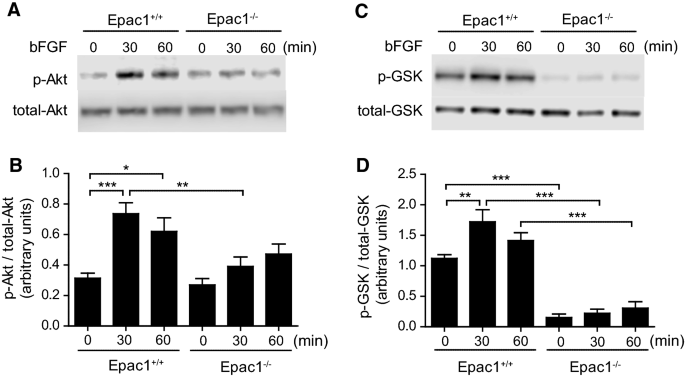
Epac1 deficiency decreased bFGF-induced phosphorylation of Akt and GSK3β. VSMCs were incubated with or without 10 ng/ml of bFGF for 30 or 60 min. A Representative western blotting images of the phosphorylation of Akt (p-Akt) are shown. B Quantification of A, n = 6. C Representative western blotting images of the phosphorylation of GSK3β (p-GSK3β) are shown. D Quantification of C, n = 6. *p < 0.05; **p < 0.01; ***p < 0.001
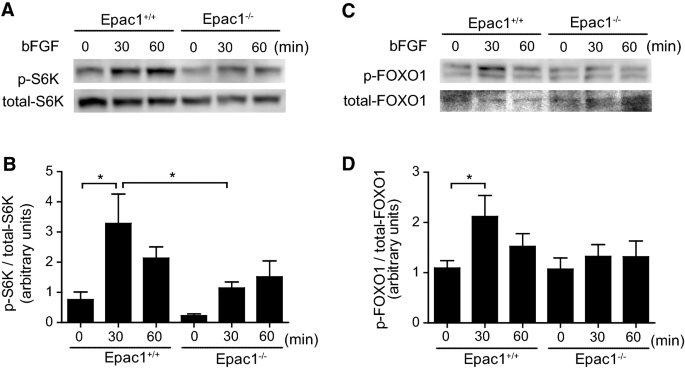
bFGF-induced phosphorylation of S6 K and FOXO1 in Epac1 deficient VSMCs. VSMCs were incubated with or without 10 ng/ml of bFGF for 30 or 60 min. A Representative western blotting images of the phosphorylation of S6 K (p-S6 K) are shown. B Quantification of A, n = 4. C Representative western blotting images of the phosphorylation of FOXO1 (p-FOXO1) are shown. D Quantification of C, n = 4. *p < 0.05
Results
Epac1 deficiency attenuated bFGF-induced VSMC migration
We examined the migratory effect of bFGF on the VSMCs of Epac1+/+ mice (Epac1+/+-VSMCs) and Epac1−/− mice (Epac1−/−-VSMCs). Cell migration was monitored for 8 h and their paths were obtained (Fig. 1a, Supplemental Movies I and II). The degree of bFGF-induced cell migration was smaller in Epac1−/−-VSMCs than in Epac1+/+-VSMCs (Fig. 1a, b). Fibroblast growth factor receptor 1 (FGFR1) mainly contributes to bFGF-induced VSMC migration [25]. Next, we examined whether FGFR1 expression was affected by Epac1 deficiency; no difference was found in FGFR1 protein expression between Epac1+/+- and Epac1−/−-VSMCs (Fig. 1c).
Epac1 deficiency decreased bFGF-induced lamellipodia formation
Lamelliopodia with membrane protrusion play fundamental roles in cell migration [26]. We next examined the effect of Epac1 deficiency on bFGF-induced lamellipodia formation and cofilin dephosphorylation, which plays a critical role in lamellipodia formation [26]. Immunocytochemistry revealed that bFGF significantly increased lamellipodia formation and was accompanied by cofilin dephosphorylation in Epac1+/+-VSMCs, but not in Epac1−/−-VSMCs (Fig. 2). These results suggest that Epac1 is involved in bFGF-induced lamellipodia formation.
Epac1 deficiency decreased bFGF-induced phosphorylation of Akt and GSK3β
It has been reported that bFGF activates PI3 K/Akt pathways, which further induces GSK3β phosphorylation, leading to cell migration [27, 28]. It is of note that PI3 K induces the phosphorylation of Akt at Ser473, thereby promoting the phosphorylation of GSK3 at Ser9, which inactivates GSK3β kinase activity [29]. We next assessed whether the phosphorylation of Akt at Ser473 and GSK3β at Ser9 are inhibited by Epac1 deficiency. bFGF stimulation increased both Akt and GSK3β phosphorylation in Epac1+/+ VSMCs, and this effect was significantly inhibited in Epac1−/− VSMCs (Fig. 3). No delayed enhancement of phosphorylation in Epac1−/− VSMCs was observed.
It has been recognized that phosphorylation of PI3 K/Akt induces S6 K and FOXO1 phosphorylation, which are related to protein synthesis and cell survival, respectively [30, 31]. bFGF stimulation increased phosphorylation of S6 K at Thr389 in Epac1+/+ VSMCs, and this effect was significantly attenuated in Epac1−/− VSMCs (Fig. 4a, b). Phosphorylation of FOXO1 at Ser256 was increased by bFGF stimulation in Epac1+/+ VSMCs. This effect was attenuated in Epac1−/− VSMCs, although this attenuation did not reach significance (Fig. 4c, d).
Epac1 deficiency suppressed intimal thickening and GSK3β phosphorylation
Based on the in vitro results, we investigated the phosphorylation status of GSK3β in mechanical injury-induced intimal thickening in Epac1+/+ and Epac1−/− mice. There was no morphological difference between the femoral arteries of Epac1+/+ or Epac1−/− mice in basal conditions (upper panels of Fig. 5). The insertion of a large wire into the femoral arteries induced prominent intimal thickening in Epac1+/+ whereas this intimal thickening was attenuated in Epac1−/− mice (lower panels of Fig. 5), which was consistent with the results of previous reports [13, 14]. Immunocytochemistry demonstrated that phosphorylated GSK3β expression was attenuated in the area of intimal thickening in Epac1−/− mice compared to that in Epac1+/+ mice (lower panels of Fig. 5).

Epac1 deficiency suppressed intimal thickening and GSK3β phosphorylation. Representative images of immunohistochemistry for GSK3β phosphorylation in the cross-sections of the uninjured (upper panels) and wire-injured (lower panels) femoral arteries of Epac1+/+ and Epac1−/− mice. Immunoreaction against GSK3β phosphorylation is shown in green. The nucleus was visualized in blue by Hoechst stain. The area between the white dotted lines indicates intimal thickening. Scale bars, 50 μm
Discussion
In the present study, we demonstrated that Epac1 deficiency attenuated the bFGF-induced phosphorylation of Akt and GSK3β, cell migration, and lamellipodia formation in VSMCs. Epac1 deficiency reduced mechanical injury-induced intimal thickening, which was accompanied by the decreased phosphorylation of GSK3β. These data indicated that Epac1-mediated downstream signaling Akt/GSK3β is involved in bFGF-induced VSMC migration.
bFGF is the major modulator of VSMC migration and intimal thickening formation. It has been demonstrated that anti-bFGF IgG administration caused an approximately 50% reduction in intimal thickening [32]. Significant inhibition of intimal thickening was observed in the artery treated with the intraluminal adenoviral gene transfer of an antisense bFGF [33]. Emerging evidence suggests that bFGF and Epac activate many cellular outcomes, including migration via PI3 K/Akt signaling [14,15,16,17,18, 20,21,22,23]. One report suggested that Epac1 increases the migration of endothelial cells via bFGF-mediated paracrine signaling [34]. In this study, Epac1 silencing inhibited the binding of bFGF to FGFR through attenuation of N-sulfation of heparin sulfate chains. The interaction between bFGF and Epac signaling pathways, however, is largely unknown.
The present study demonstrated that the bFGF-mediated phosphorylation of Akt and GSK3β and the migration in VSMCs were significantly attenuated under the Epac1-deficient condition, suggesting an association between Epac1- and bFGF-mediated downstream signaling. Epac1 is activated by direct cAMP binding and mediates a wide range of cellular responses [35, 36]. A previous paper proposed the mechanisms of bFGF stimulation-mediated cAMP production [37]. FGFR1 activates the PLCγ/diacylglycerol pathway, enhancing Ca2+ entry through voltage-gated calcium channels, and Ca2+/calmodulin could then activate calcium-sensitive adenylyl cyclases, producing cAMP [37]. Although this paper proposed this signaling cascade, the concept was not supported by the experimental data. It is, however, recognized that bFGF is not directly involved in the elevation of the intracellular concentration of cAMP [38,39,40]. Based on these findings, there is currently no evidence that bFGF-FGFR signaling directly regulates Epac1 activation. We assume that the bFGF-mediated downstream signaling interacts with the Epac1-mediated intracellular signaling pathways. We do not, however, know the exact molecular mechanisms of this interaction. Further study on small G protein Rap and PI3 K is needed to determine how the bFGF and Epac1 signaling pathways interact.
In the present study, we found low levels of GSK3β phosphorylation in Epac1−/−-VSMCs with or without bFGF stimulation. GSK3β ubiquitously expresses serine/threonine kinase and has a high basal activity in resting cells. GSK3β plays roles in many biological processes, including cell polarity, microtubule dynamics, and cell migration [41,42,43]. The importance of GSK3β inactivation in vascular remodeling has also been reported [44]. In a rat balloon catheter injury model, the expression of phosphorylated GSK3β (inactivation) was markedly increased in the intimal thickening area, and the constitutively active GSK3β gene transfer resulted in reduced intimal thickening formation [44]. During intimal thickening formation, VSMC apoptosis was inhibited by the upregulation of glucose metabolism and is linked to the inactivation of GSK3β [45].
The role of Epac in GSK3β activation has been controversial. Studies using skeletal muscle cells, hippocampal slices, and cultured microglia demonstrated no effect of Epac on GSK3β activation [46,47,48]. Two papers, however, showed that Epac or Epac1 inactivated GSK3β in isolated rat heart and melanoma cells [27, 49]. Akt is a major upstream regulator that results in the inactivation of GSK3β through the phosphorylation of the N-terminal Ser9 [29]. Epac promoted tubulin polymerization via activation of the PI3 K/Akt/GSK3β pathway [27]. These data suggest that Epac-mediated GSK3β activation depends on the cell type and upstream of Epac signaling. Our data showed that, under basal conditions, the degree of phosphorylation of Akt was similar between Epac1+/+- and Epac1−/−-VSMCs, whereas the expression level of phosphorylated GSK3β was lower in Epac1−/−-VSMCs than in Epac1+/+-VSMCs. The mitogen-activated protein kinase cascade and the Wnt pathway are the upstream signaling of GSK3β, as well as the PI3 K/Akt axis [50, 51]. In addition to Akt activation, Epac-mediated upstream signaling that activates GSK3β may exist.
GSK3β inactivation leads to activation of the β-catenin signaling pathway via the accumulation of β-catenin in the nucleus, which regulates gene expression and a variety of cell activities, including cell migration [52]. It has been reported that bFGF promotes skin fibroblast migration via the GSK3β/β-catenin signaling pathway [15]. An elevated level of active β-catenin was detected at the sites of intimal thickening after vascular injury, and inhibition of β-catenin decreased neointima formation after injury [53]. These data suggest the possible role of β-catenin in VSMC migration. The mechanisms connecting GSK3β inactivation and VSMC migration need to be determined in a future study.
In addition to cell migration, Akt signaling regulates protein synthesis and cell survival or proliferation via Akt/S6 K and Akt/FOXO1 signaling pathways, respectively [54]. Our data suggested that S6 K-mediated protein synthesis and FOXO1-mediated cell survival or proliferation were attenuated in Epac1−/−-VSMCs. Although a few papers indicated the involvement of S6 K and FOXO1 in intimal thickening formation [55, 56] and there is currently no direct evidence demonstrating causal relationship between these signaling pathways and vascular intimal thickening formation, Epac1-mediated regulation of these signaling pathways would be important for a future study.
In conclusion, Epac1 plays a role in bFGF-induced VSMC migration via Akt/GSK3β phosphorylation. Both our present study and our previous study [13] suggest that Epac1 is involved in PDGF-BB and bFGF-mediated VSMC migration. It has been reported that Epac1 was upregulated during injury-induced intimal thickening formation [10] and that Epac1 deficiency or the pharmacological inhibition of Epac attenuated intimal thickening formation in a mouse model of vascular injury [13, 14]. Epac1 may be a future target for therapy to inhibit vascular intimal thickening.
References
Schwartz SM (1997) Perspectives series: cell adhesion in vascular biology. Smooth muscle migration in atherosclerosis and restenosis. J Clin Investig 99:2814–2816
Newby AC, Zaltsman AB (2000) Molecular mechanisms in intimal hyperplasia. J Pathol 190:300–309
Stolker JM, Kennedy KF, Lindsey JB, Marso SP, Pencina MJ, Cutlip DE, Mauri L, Kleiman NS, Cohen DJ, Investigators E (2010) Predicting restenosis of drug-eluting stents placed in real-world clinical practice: derivation and validation of a risk model from the EVENT registry. Circ Cardiovasc Interv 3:327–334
Finn AV, Joner M, Nakazawa G, Kolodgie F, Newell J, John MC, Gold HK, Virmani R (2007) Pathological correlates of late drug-eluting stent thrombosis: strut coverage as a marker of endothelialization. Circulation 115:2435–2441
Casscells W (1992) Migration of smooth muscle and endothelial cells. Critical events in restenosis. Circulation 86:723–729
Jackson CL, Reidy MA (1993) Basic fibroblast growth factor: its role in the control of smooth muscle cell migration. Am J Pathol 143:1024–1031
Schroder K, Helmcke I, Palfi K, Krause KH, Busse R, Brandes RP (2007) Nox1 mediates basic fibroblast growth factor-induced migration of vascular smooth muscle cells. Arterioscler Thromb Vasc Biol 27:1736–1743
Lindner V, Lappi DA, Baird A, Majack RA, Reidy MA (1991) Role of basic fibroblast growth factor in vascular lesion formation. Circ Res 68:106–113
Lindner V (1995) Role of basic fibroblast growth factor and platelet-derived growth factor (B-chain) in neointima formation after arterial injury. Z Kardiol 84(Suppl 4):137–144
Yokoyama U, Minamisawa S, Quan H, Akaike T, Jin M, Otsu K, Ulucan C, Wang X, Baljinnyam E, Takaoka M, Sata M, Ishikawa Y (2008) Epac1 is upregulated during neointima formation and promotes vascular smooth muscle cell migration. Am J Physiol Heart Circ Physiol 295:H1547–H1555
Yokoyama U, Minamisawa S, Quan H, Akaike T, Suzuki S, Jin M, Jiao Q, Watanabe M, Otsu K, Iwasaki S, Nishimaki S, Sato M, Ishikawa Y (2008) Prostaglandin E2-activated Epac promotes neointimal formation of the rat ductus arteriosus by a process distinct from that of cAMP-dependent protein kinase A. J Biol Chem 283:28702–28709
Yokoyama U, Patel HH, Lai NC, Aroonsakool N, Roth DM, Insel PA (2008) The cyclic AMP effector Epac integrates pro- and anti-fibrotic signals. Proc Natl Acad Sci USA 105:6386–6391
Kato Y, Yokoyama U, Yanai C, Ishige R, Kurotaki D, Umemura M, Fujita T, Kubota T, Okumura S, Sata M, Tamura T, Ishikawa Y (2015) Epac1 deficiency attenuated vascular smooth muscle cell migration and neointimal formation. Arterioscler Thromb Vasc Biol 35:2617–2625
Wang H, Robichaux WG, Wang Z, Mei FC, Cai M, Du G, Chen J, Cheng X (2016) Inhibition of Epac1 suppresses mitochondrial fission and reduces neointima formation induced by vascular injury. Sci Rep 6:36552
Wang X, Zhu Y, Sun C, Wang T, Shen Y, Cai W, Sun J, Chi L, Wang H, Song N, Niu C, Shen J, Cong W, Zhu Z, Xuan Y, Li X, Jin L (2017) Feedback activation of basic fibroblast growth factor signaling via the Wnt/beta-catenin pathway in skin fibroblasts. Front Pharmacol 8:32
Schmidt A, Ladage D, Schinkothe T, Klausmann U, Ulrichs C, Klinz FJ, Brixius K, Arnhold S, Desai B, Mehlhorn U, Schwinger RH, Staib P, Addicks K, Bloch W (2006) Basic fibroblast growth factor controls migration in human mesenchymal stem cells. Stem Cells 24:1750–1758
Shi H, Lin B, Huang Y, Wu J, Zhang H, Lin C, Wang Z, Zhu J, Zhao Y, Fu X, Lou Z, Li X, Xiao J (2016) Basic fibroblast growth factor promotes melanocyte migration via activating PI3 K/Akt-Rac1-FAK-JNK and ERK signaling pathways. IUBMB Life 68:735–747
Shimabukuro Y, Terashima H, Takedachi M, Maeda K, Nakamura T, Sawada K, Kobashi M, Awata T, Oohara H, Kawahara T, Iwayama T, Hashikawa T, Yanagita M, Yamada S, Murakami S (2011) Fibroblast growth factor-2 stimulates directed migration of periodontal ligament cells via PI3 K/AKT signaling and CD44/hyaluronan interaction. J Cell Physiol 226:809–821
Shigematsu K, Koyama H, Olson NE, Cho A, Reidy MA (2000) Phosphatidylinositol 3-kinase signaling is important for smooth muscle cell replication after arterial injury. Arterioscler Thromb Vasc Biol 20:2373–2378
Jing H, Yen JH, Ganea D (2004) A novel signaling pathway mediates the inhibition of CCL3/4 expression by prostaglandin E2. J Biol Chem 279:55176–55186
Namkoong S, Kim CK, Cho YL, Kim JH, Lee H, Ha KS, Choe J, Kim PH, Won MH, Kwon YG, Shim EB, Kim YM (2009) Forskolin increases angiogenesis through the coordinated cross-talk of PKA-dependent VEGF expression and Epac-mediated PI3 K/Akt/eNOS signaling. Cell Signal 21:906–915
Villarreal F, Epperson SA, Ramirez-Sanchez I, Yamazaki KG, Brunton LL (2009) Regulation of cardiac fibroblast collagen synthesis by adenosine: roles for Epac and PI3 K. Am J Physiol Cell Physiol 296:C1178–C1184
Baviera AM, Zanon NM, Navegantes LC, Kettelhut IC (2010) Involvement of cAMP/Epac/PI3K-dependent pathway in the antiproteolytic effect of epinephrine on rat skeletal muscle. Mol Cell Endocrinol 315:104–112
Suzuki S, Yokoyama U, Abe T, Kiyonari H, Yamashita N, Kato Y, Kurotani R, Sato M, Okumura S, Ishikawa Y (2010) Differential roles of Epac in regulating cell death in neuronal and myocardial cells. J Biol Chem 285:24248–24259
Rauch BH, Millette E, Kenagy RD, Daum G, Fischer JW, Clowes AW (2005) Syndecan-4 is required for thrombin-induced migration and proliferation in human vascular smooth muscle cells. J Biol Chem 280:17507–17511
Ghosh M, Song X, Mouneimne G, Sidani M, Lawrence DS, Condeelis JS (2004) Cofilin promotes actin polymerization and defines the direction of cell motility. Science 304:743–746
Baljinnyam E, Iwatsubo K, Kurotani R, Wang X, Ulucan C, Iwatsubo M, Lagunoff D, Ishikawa Y (2009) Epac increases melanoma cell migration by a heparan sulfate-related mechanism. Am J Physiol Cell Physiol 297:C802–C813
Liu ZC, Wang HS, Zhang G, Liu H, Chen XH, Zhang F, Chen DY, Cai SH, Du J (2014) AKT/GSK-3beta regulates stability and transcription of snail which is crucial for bFGF-induced epithelial-mesenchymal transition of prostate cancer cells. Biochim Biophys Acta 1840:3096–3105
Cohen P, Frame S (2001) The renaissance of GSK3. Nat Rev Mol Cell Biol 2:769–776
Pullen N, Thomas G (1997) The modular phosphorylation and activation of p70s6k. FEBS Lett 410:78–82
Huang H, Tindall DJ (2007) Dynamic FoxO transcription factors. J Cell Sci 120:2479–2487
Rutherford C, Martin W, Salame M, Carrier M, Anggard E, Ferns G (1997) Substantial inhibition of neo-intimal response to balloon injury in the rat carotid artery using a combination of antibodies to platelet-derived growth factor-BB and basic fibroblast growth factor. Atherosclerosis 130:45–51
Hanna AK, Fox JC, Neschis DG, Safford SD, Swain JL, Golden MA (1997) Antisense basic fibroblast growth factor gene transfer reduces neointimal thickening after arterial injury. J Vasc Surg 25:320–325
Baljinnyam E, Umemura M, Chuang C, De Lorenzo MS, Iwatsubo M, Chen S, Goydos JS, Ishikawa Y, Whitelock JM, Iwatsubo K (2014) Epac1 increases migration of endothelial cells and melanoma cells via FGF2-mediated paracrine signaling. Pigment Cell Melanoma Res 27:611–620
de Rooij J, Zwartkruis FJ, Verheijen MH, Cool RH, Nijman SM, Wittinghofer A, Bos JL (1998) Epac is a Rap1 guanine-nucleotide-exchange factor directly activated by cyclic AMP. Nature 396:474–477
Gloerich M, Bos JL (2010) Epac: defining a new mechanism for cAMP action. Annu Rev Pharmacol Toxicol 50:355–375
Soto I, Rosenthal JJ, Blagburn JM, Blanco RE (2006) Fibroblast growth factor 2 applied to the optic nerve after axotomy up-regulates BDNF and TrkB in ganglion cells by activating the ERK and PKA signaling pathways. J Neurochem 96:82–96
Pursiheimo JP, Jalkanen M, Tasken K, Jaakkola P (2000) Involvement of protein kinase A in fibroblast growth factor-2-activated transcription. Proc Natl Acad Sci USA 97:168–173
Yoshimata T, Yoneyama A, Jin-no Y, Tamai N, Kamiya Y (1999) Effects of dehydroepiandrosterone on mitogen-activated protein kinase in human aortic smooth muscle cells. Life Sci 65:431–440
Logan A, Logan SD (1991) Studies on the mechanisms of signalling and inhibition by pertussis toxin of fibroblast growth factor-stimulated mitogenesis in Balb/c 3T3 cells. Cell Signal 3:215–223
Jiang H, Guo W, Liang X, Rao Y (2005) Both the establishment and the maintenance of neuronal polarity require active mechanisms: critical roles of GSK-3beta and its upstream regulators. Cell 120:123–135
Doble BW, Woodgett JR (2003) GSK-3: tricks of the trade for a multi-tasking kinase. J Cell Sci 116:1175–1186
Etienne-Manneville S, Hall A (2003) Cdc42 regulates GSK-3beta and adenomatous polyposis coli to control cell polarity. Nature 421:753–756
Park KW, Yang HM, Youn SW, Yang HJ, Chae IH, Oh BH, Lee MM, Park YB, Choi YS, Kim HS, Walsh K (2003) Constitutively active glycogen synthase kinase-3beta gene transfer sustains apoptosis, inhibits proliferation of vascular smooth muscle cells, and reduces neointima formation after balloon injury in rats. Arterioscler Thromb Vasc Biol 23:1364–1369
Hall JL, Chatham JC, Eldar-Finkelman H, Gibbons GH (2001) Upregulation of glucose metabolism during intimal lesion formation is coupled to the inhibition of vascular smooth muscle cell apoptosis. Role of GSK3beta. Diabetes 50:1171–1179
Jensen J, Brennesvik EO, Lai YC, Shepherd PR (2007) GSK-3beta regulation in skeletal muscles by adrenaline and insulin: evidence that PKA and PKB regulate different pools of GSK-3. Cell Signal 19:204–210
Ster J, de Bock F, Bertaso F, Abitbol K, Daniel H, Bockaert J, Fagni L (2009) Epac mediates PACAP-dependent long-term depression in the hippocampus. J Physiol 587:101–113
Liu J, Zhao X, Cao J, Xue Q, Feng X, Liu X, Zhang F, Yu B (2011) Differential roles of PKA and Epac on the production of cytokines in the endotoxin-stimulated primary cultured microglia. J Mol Neurosci MN 45:186–193
Lee TM, Lin SZ, Chang NC (2013) Both PKA and Epac pathways mediate N-acetylcysteine-induced Connexin43 preservation in rats with myocardial infarction. PLoS One 8:e71878
Sutherland C, Leighton IA, Cohen P (1993) Inactivation of glycogen synthase kinase-3 beta by phosphorylation: new kinase connections in insulin and growth-factor signalling. Biochem J 296(Pt 1):15–19
Cook D, Fry MJ, Hughes K, Sumathipala R, Woodgett JR, Dale TC (1996) Wingless inactivates glycogen synthase kinase-3 via an intracellular signalling pathway which involves a protein kinase C. EMBO J 15:4526–4536
Gordon MD, Nusse R (2006) Wnt signaling: multiple pathways, multiple receptors, and multiple transcription factors. J Biol Chem 281:22429–22433
Riascos-Bernal DF, Chinnasamy P, Gross JN, Almonte V, Egana-Gorrono L, Parikh D, Jayakumar S, Guo L, Sibinga NES (2017) Inhibition of smooth muscle beta-catenin hinders neointima formation after vascular injury. Arterioscler Thromb Vasc Biol 37:879–888
Manning BD, Cantley LC (2007) AKT/PKB signaling: navigating downstream. Cell 129:1261–1274
Zhu LH, Huang L, Zhang X, Zhang P, Zhang SM, Guan H, Zhang Y, Zhu XY, Tian S, Deng K, Li H (2015) Mindin regulates vascular smooth muscle cell phenotype and prevents neointima formation. Clin Sci 129:129–145
Wang Y, Bai Y, Qin L, Zhang P, Yi T, Teesdale SA, Zhao L, Pober JS, Tellides G (2007) Interferon-gamma induces human vascular smooth muscle cell proliferation and intimal expansion by phosphatidylinositol 3-kinase dependent mammalian target of rapamycin raptor complex 1 activation. Circ Res 101:560–569
Acknowledgements
The authors are grateful to Yuka Sawada (Yokohama City University) for histological analysis.
Funding
This study was funded by MEXT/JSPS KAKENHI (YK, JP17K08976; UY, JP17K19403, JP16H05358, JP15H05761; YI, JPH1605300), the Takeda Science Foundation (YK), the Japan Agency for Medical Research and Development (AMED) (YI, 66890007, 66891153), and the Kitsuen Research Foundation (YI, 71890005).
Author information
Authors and Affiliations
Corresponding authors
Electronic supplementary material
Below is the link to the electronic supplementary material.
Supplemental movie I. Epac1+/+-VSMC migration under stimulation with 10 ng/ml of bFGF. (MPG 887 kb)
Supplemental movie II. Epac1−/−-VSMC migration under stimulation with 10 ng/ml of bFGF. (MPG 885 kb)
Rights and permissions
Springer Nature or its licensor (e.g. a society or other partner) holds exclusive rights to this article under a publishing agreement with the author(s) or other rightsholder(s); author self-archiving of the accepted manuscript version of this article is solely governed by the terms of such publishing agreement and applicable law.
About this article

Cite this article
Kato, Y., Yokoyama, U., Fujita, T. et al. Epac1 deficiency inhibits basic fibroblast growth factor-mediated vascular smooth muscle cell migration. J Physiol Sci 69, 175–184 (2019). https://doi.org/10.1007/s12576-018-0631-7
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s12576-018-0631-7